
细菌糖原分支酶是如何进化的?
糖原是一种多分枝多糖(糖),用于动物、真菌和细菌的能量储存。它是一种非常有用的快速和容易获取葡萄糖和能量的来源。在细菌中,它在储存碳和能量方面起着重要作用。
细菌是难以置信的顽强生物,能够在地球上一些最极端的环境中生存,比如深海喷口。细菌能够在不利时期分解并利用糖原作为能量来源,这有助于提高它们长期生存的机会。
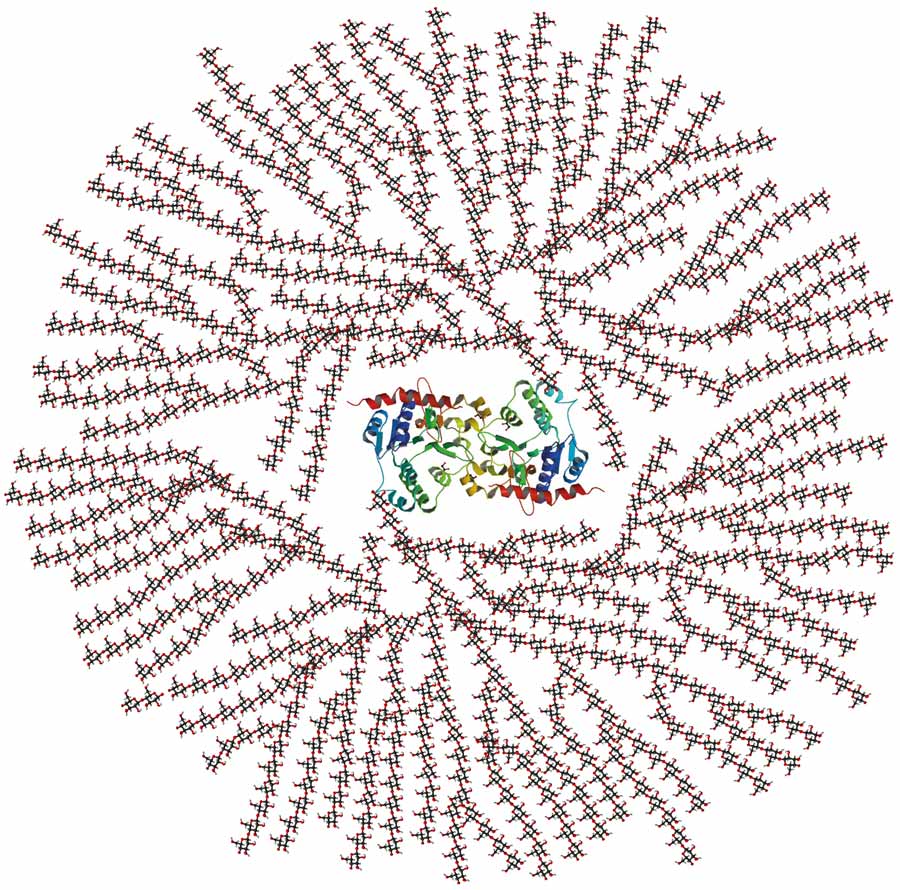
细菌酶和糖原
细菌中有大量的酶与糖原的功能有关。其中,glgB- 额定支化酶,否则被称为GBE,毫无疑问是最重要的一个。GBE是细菌糖原代谢的重要基因之一,是糖原结构的决定因素。
的GBE基因在形成α-1,6-糖苷分支点中起着至关重要的作用,决定着糖原中独特的分支模式。正是这些分支使得它们在生物体中很容易获得能量。
细菌可以在不利时期分解并利用糖原作为能量来源,这有助于提高它们长期生存的机会。
上海巴斯德研究所的王亮博士和澳门科技大学的刘庆华女士试图理解GBE通过对其结构和进化的研究来更好地研究基因。糖原结构与人类2型糖尿病的血糖控制有关,因此更清楚地了解细菌中糖原结构的调节,也可能有助于了解高等生物中糖原结构的异常。
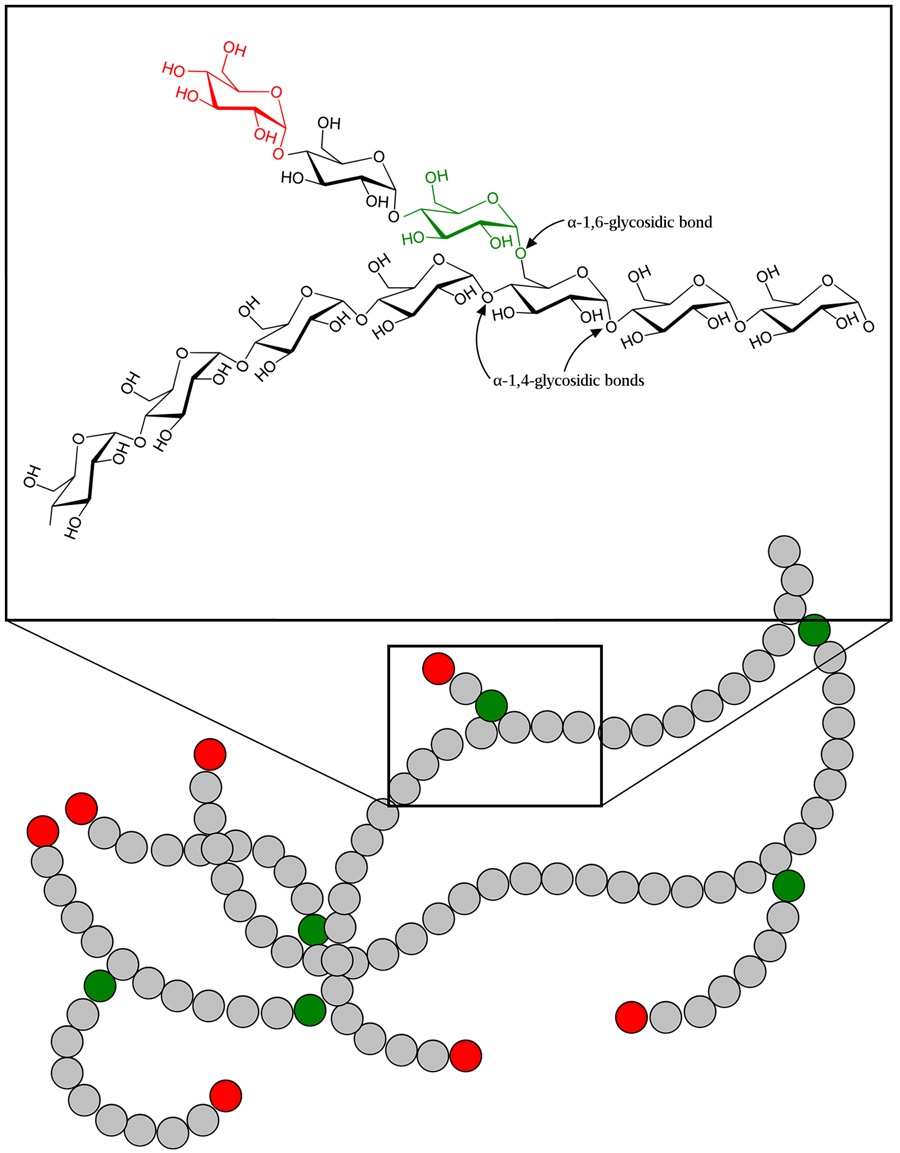
GBEs是如何进化的?
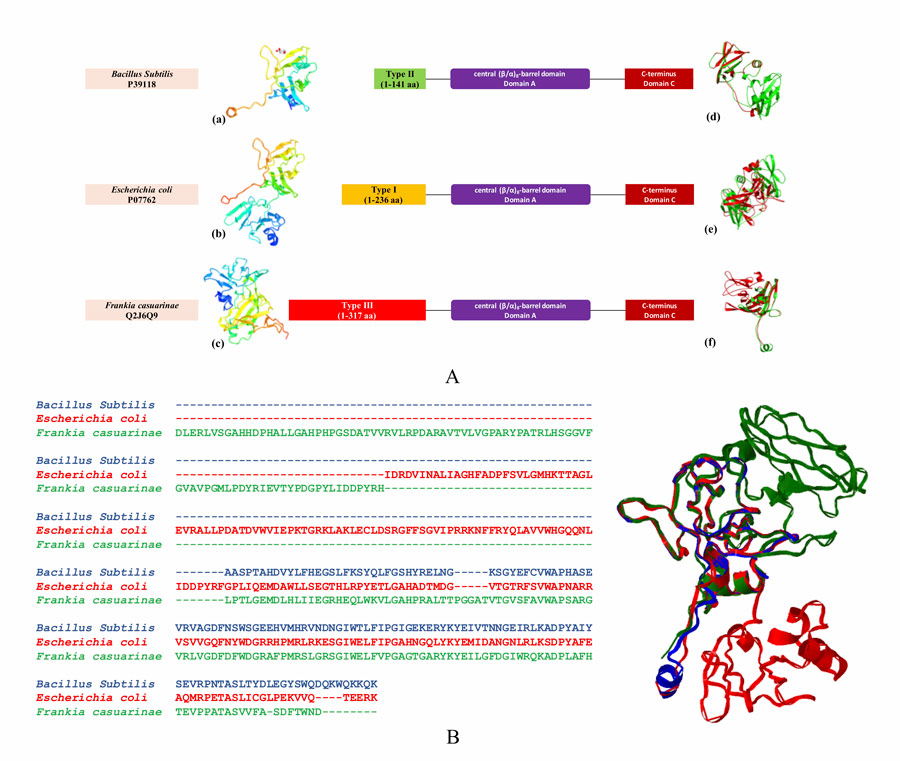
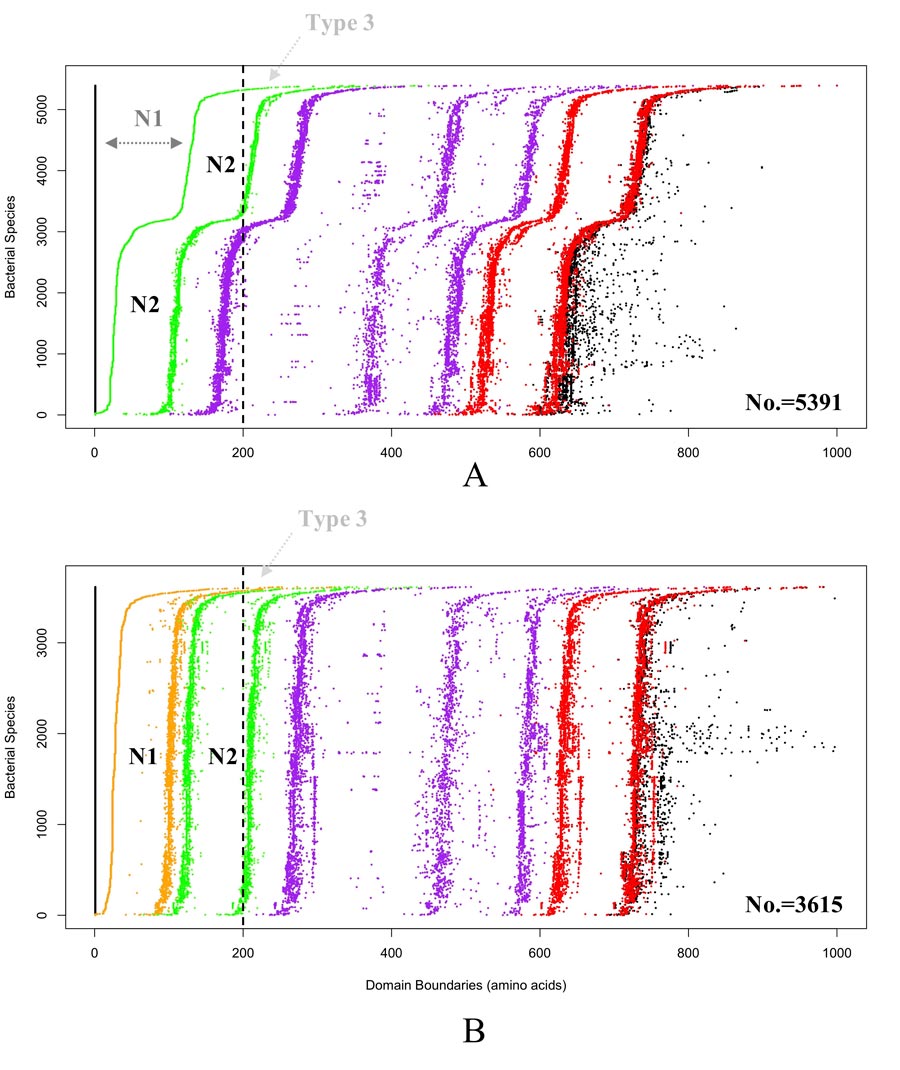
以前对一小组gbe的演化分析表明,可能存在两种类型的gbe。这是基于它们的n端结构域。n端是位于形成蛋白质的多肽链起始位置的胺基(-NH2)。第一种类型的GBE(类型1)有N1和N2(即CBM48)域,而第二种类型的GBE(类型2)只有N2域。
在Wang和Liu的研究中,他们基于隐马尔可夫模型初步分析了169个人工审查的细菌GBEs的n端结构域。在概率论中,马尔可夫模型被用来模拟随机变化的系统。当状态是部分可观测的时候,使用隐马尔可夫模型。
研究期间的系统发育分析发现了某些细菌植物中的GBE类型的聚类模式(王国低于王国的生物分类水平)。该分析显示,较短的2种GBE主要以革兰氏阳性物种发现,而在革兰氏阴性物质中主要发现1种GBE的较长型。
革兰氏阳性和革兰氏阴性描述了细菌外层的差异。革兰氏阳性细菌有厚厚的肽聚糖层,可以吸收周围的物质,而革兰氏阴性细菌有多层薄膜层,可以排泄毒素。
有趣的是,该研究还发现了此前未报道的第三组GBE (Type 3)。这些Type 3 GBE在N1结构域之前有大约100个氨基酸。
这对细菌代谢意味着什么?
过去的几项体外研究已经将N1结构域与糖原形成过程中短寡糖(糖聚合物,通常含有3到10个单糖)链的转移联系起来。这反过来又会导致小而紧凑的糖原结构。
这些紧凑的糖原结构更慢地降解。因此,它们可以作为特别耐用的能量储备,这可能有助于细菌所显示的增强的环境持久性。
研究人员认为,这可能形成基于其n端结构域的GBEs新分类的基础。他们进行了大规模的序列分析,看看是否能找到任何模式。他们的目标是通过在物种和序列水平上的系统发育分析(研究一个物种或一组生物或一个生物的特定特征的进化发展)来了解不同类型GBEs的进化模式。研究人员还对GBE的n端进行了三维建模,以提供结构比较。
这可能是潜在的服务
作为酶促操纵糖原结构的生物技术工具。
研究结果
研究人员从序列演化和结构域的角度对数千种细菌中的GBEs进行了系统研究。他们对9387个GBE序列进行了分析。该分析确定了147个GBE,它们可能属于3型GBE的一个新的(可能是新的)组。这些细菌大部分属于放线菌门(一群可以生活在陆地或水中的革兰氏阳性细菌)。它们的DNA中鸟嘌呤和胞嘧啶含量高,没有明显的细胞壁)。
他们还研究了不同GBE类型中糖原链的平均长度之间是否存在相关性。不幸的是,由于数据有限,在这方面的研究无法得出重要的结论。
参考
1.Abad MC, Binderup K, Rios-Steiner J, Arni RK, Preiss J, Geiger JH:大肠杆菌分支酶的x射线晶体结构.中国生物医学工程学报,2003,23(4):491 - 498。
2. Waterhouse A,Bertoni M,Bienert S,Studer G,Tauriello G,Gumienny R,Heer Ft,De Beer Ta P,Rempffer C,Bordoli L等人:SWISS-MODEL:蛋白质结构和复合物的同源建模.核酸研究2018,46 (W1):W296-W303。
虽然观察到一些差异,但本研究相关性分析显示GBE类型与糖原一级结构之间无统计学意义。对不相关的一种可能解释是,该分析从各种来源获得的数据量不足。
未来的前景
由王博士领导的团队认为,未来的研究将受益于将标准化的糖原提取和结构表征技术应用于更多的细菌物种。他们还认为,考虑到GBE n端对糖原结构的重要性,研究3型GBE中扩展n端的功能,并与1型和2型GBE的结果进行比较,肯定是值得的。
该团队还建议在模型微生物中建立三种GBE的原位表达,如大肠杆菌和比较糖原细分子结构可以提示不同类型的N-末端与糖原初级结构相关的暗示。
值得注意的是,细菌GBE中的N-末端结构域具有高变异性和比较唯一性。这可能是作为抗生素治疗或生物技术工具的未来药物,用于酶促操纵糖原结构。令人兴奋的是,未来的治疗可以通过糖原结构与细菌的能源之间的潜在联系来帮助减弱细菌持久性。
个人反应
在你发现可能存在第三种细菌GBE之后,你认为最有趣的应用是什么?
在真核生物和原核生物中,糖原颗粒的精细分子结构与其生理功能密切相关,如血糖稳态和渗透平衡等。GBE部分控制糖原的初级结构,即链长分布,它影响糖原颗粒大小、分支点百分比和降解率。前人研究证实GBEs的长n端与短寡糖的转移有关。因此,在糖原合成过程中,具有较长n端的新型第三类GBEs能够转移较短的链,这可能是一种控制糖原结构的生物技术工具,可用于工业应用。