
新的RNA基因是如何产生的
基因复制,即从现有基因的精确复制中形成新的基因,长期以来一直被认为是基因形成背后的主要过程。然而,已有研究表明,基因可能来自非编码DNA,即具有开放阅读框并显示翻译活性但不编码蛋白质的DNA区域。这一过程被称为基于原基因序列或不代表现有基因的序列的新生蛋白基因形成。这一概念是由匹兹堡大学的Anne-Ruxandra Carvunis博士及其同事以及其他机构的研究人员提出的。它构成了一个模型来解释在进化过程中,新的和不同的蛋白质基因是如何通过从头生成机制而不是现有的基因序列复制产生的。

纽约石溪大学文艺复兴医学院的Nicholas Delihas博士最初试图了解RNA基因家族中的重复DNA序列,即在人类第22号染色体中发现的FAM230长基因间非编码RNA (lincRNA)家族。这种重复序列和RNA基因家族可能与人类遗传疾病的发病有关,包括异常的染色体重组和随后的染色体缺失。然而,这些研究导致一个意想不到的发现的祖先DNA重复序列,称为FAM247,这可以作为一个gene-forming元素:成新的RNA基因开发相似的概念设想新创蛋白质Carvunis博士出生的同事和其他机构的科学家们。
一种进化的保守序列形成了各种基因的一部分,并且似乎是在进化期间开发新基因的成核位点。
进化科学家经常调查人类和灵长类以及其他物种的遗传结构,以发现不同基因在不同种群适应环境的方式中发挥的具体作用,并绘制出在进化过程中基因组发生的任何变化。最近,人们主要关注长非编码RNA (lncrna)基因的基因组测定和测序,这些基因现在被认为是许多生化途径的关键角色。这就引出了一个问题:基因组中有多少lncRNA基因,它们在进化过程中的年龄以及它们是如何出生的。
随着对De Novo基因的研究的兴趣增加,即由划痕而不是来自模板的基因,这也导致了关于事实构成基因的问题。通常同意遗传序列是导致形成功能性产品的遗传序列,其可能是RNA或蛋白质。存在不同的方法,以确认基因是功能实体。一种方法是通过生物化学技术确认RNA和蛋白质水平的基因表达。另一种方法是破坏特定的遗传序列并观察表型中发生任何变化。然而,这可能在分析整个基因组时是有问题的。进化方法看起来有特定的遗传'签名',其提供选择的证据,试图确认基因的存在。另一方面,尽管与基因的鉴定相关的挑战,但现在有足够的证据表明在整个进化历史上发生了De Novo基因形成的现象。
发现进化保守的成核位点
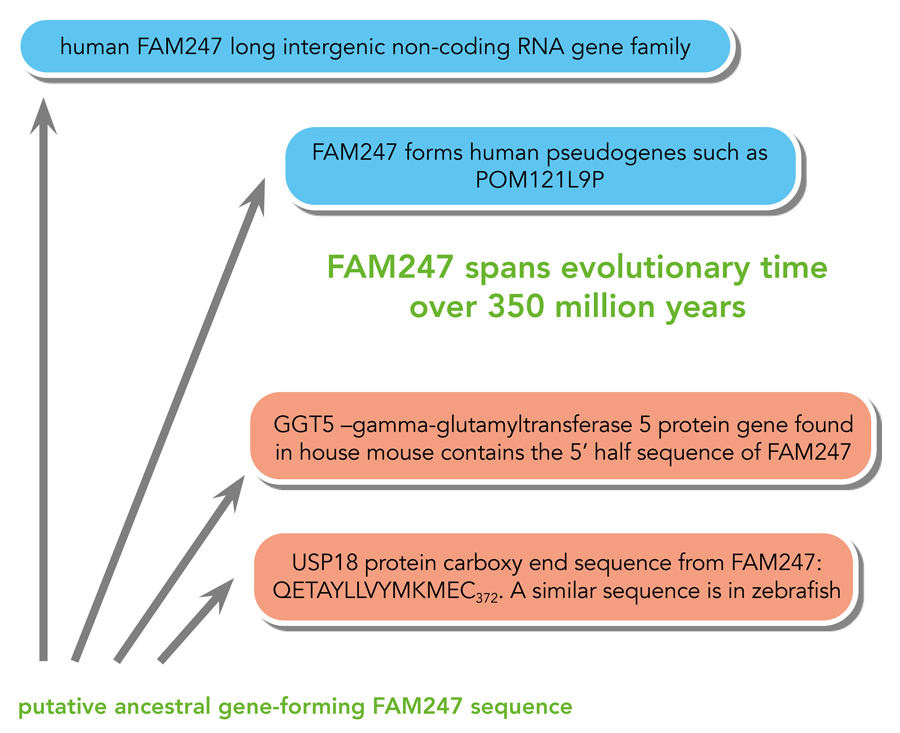
最近在人类中形成了5个FAM247 lincRNA家族基因,该基因家族似乎起源于FAM247序列的基因序列复制,该序列长度为11231 bp。这与通过基因复制建立的基因家族起源模型一致。然而,其他基因、假基因和蛋白基因也发现含有FAM247 lincRNA家族序列片段(见图1).这些伪基因特别有趣,因为它们是独特的,包含与FAM247或起源的亲本基因无关的外源染色体序列;因此,它们与真正的伪基因有很大的不同。伪基因是指具有蛋白质基因序列的副本,但由于序列中的关键突变而不能形成蛋白质产物的基因。

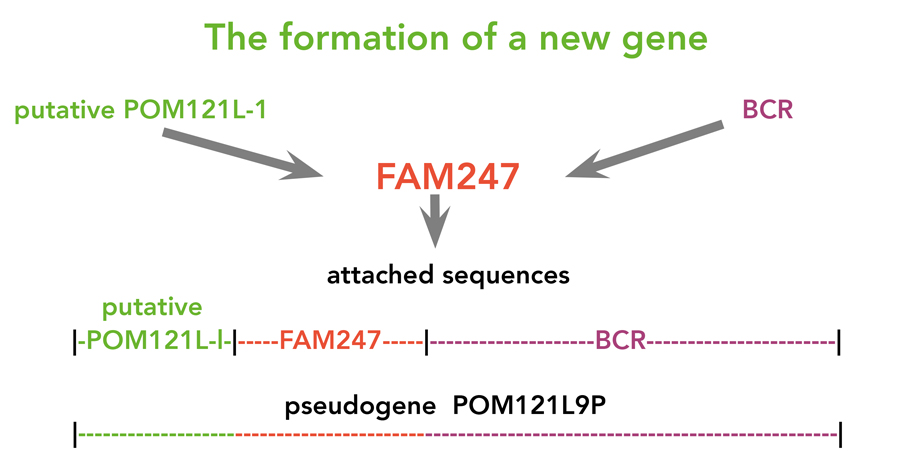
Delihas博士提出了一个包含这些所谓伪基因的基因形成模型,其中FAM247片段作为成核元素或基础位点。来自基因组其他部分的其他序列块被添加到FAM247序列中,形成成熟基因(见图2).这些FAM247片段在基因组区域中,显示出进化保守的序列签名。FAM247序列似乎携带这些无关序列附着的信息,并且它代表了该和其他伪原的DE Novo Genesis的焦点。这些假生素显示RNA转录物表达,在各种组织中以非常宽且稳健的方式显示几种。这些基因的功能尚不清楚。然而,基因函数的测定是评估这些伪原与细胞分子过程的相关程度。


令人惊讶的是,两个古老的蛋白质基因,γ -谷氨酰基转移酶(GGT5)和泛素特异性肽酶18 USP18也发现了FAM247序列片段(见图1).这两种基因都可以追溯到一亿到几亿年前的进化史。因此,在脊椎动物的进化过程中,FAM247序列形成了多种基因的一部分。不同于从头形成的修饰伪基因和由基因复制形成的5个人类FAM247 lincRNA家族基因,导致脊椎动物祖先中蛋白质基因形成的机制尚不清楚。
该小型基因组DNA序列似乎携带用于附着其他基因组区段的信息,这导致新基因的形成。它还在其开放阅读框架中携带蛋白质外显子的信息。
FAM247提供的进化高度保守序列对于免疫系统的调节至关重要
这些研究的一个主要亮点是FAM247在提供USP18基因在免疫系统调节中所必需的氨基酸序列方面的重要作用。USP18是泛素特异性肽酶基因,是去泛素化蛋白酶家族的成员。许多蛋白质通过泛素的加入而被修饰。当它附着在蛋白质上时,它就像一个信号分子,控制细胞的分化,并在免疫系统的调节中发挥作用。去泛素化蛋白酶,如USP18,通过关闭泛素化后获得的信号,从蛋白质中去除泛素,调节其功能。

FAM247核苷酸序列与USP18的人类/灵长类的羧基末端外显子核苷酸序列具有很高的同源性,大多数情况下同源性为100%。在斑马鱼中,氨基酸序列对比显示,与人类USP18末端外显子和FAM247翻译序列相比,末端外显子的少量氨基酸发生了变化,但大部分氨基酸未发生变化。首先,在斑马鱼USP18基因的末端外显子中存在大量的系统保守氨基酸位置,表明FAM247序列或其部分序列存在于斑马鱼USP18基因中。无论对蛋白质功能多么重要,最后一个外显子的大部分氨基酸序列在进化上是保守的,这可能意味着泛素特异性肽酶USP18的某些功能起源于数亿年前的脊椎动物。在一系列的论文中,加州大学圣地亚哥分校的Dong-Er Zhang博士和其他机构的同事报道了USP18蛋白羧基端功能的阐明。这些功能具有重要意义,因为它们对干扰素的调节有影响。干扰素是在应对病毒感染时释放的,并负责炎症级联反应,增强抗病毒防御。USP18是干扰素信号的负调控因子,作为一种抗炎蛋白。此外,USP18在病原控制和自身免疫性疾病的发展中具有重要作用。因此,该蛋白可以作为与炎症相关的各种疾病的重要药理学靶点。

个人反应
你目前是在关注22号染色体的重复序列还是有其他的计划?
我目前正专注于在22号染色体上寻找可能作为原基因元件的额外重复序列